For almost any biological process, seeing is believing. Because of the role viruses play in infectious disease, many biologists would like to observe them doing their thing in real time. Fluorescence imaging provides one of the few noninvasive ways to observe how viruses transport their contents into a cell and how new viruses are assembled. Such studies require high-quality microscopes with sensitive cameras that can shoot multiple frames per second. As with cells, researchers have to make sure that the position of fluorescent labels and the experimental setup do not interfere with the behavior of the system that they are studying.
Viruses are small, and they can be structurally and functionally complex. While fluorescence imaging offers the unique opportunity to observe viruses both directly and noninvasively, there are plenty of pitfalls, says Börries Brandenburg, a senior scientist at Crucell, a biotechnology company in Leiden, Netherlands. “You need...
The Scientist sought out the experts to bring you a guide on imaging viral behavior in living systems.
Choosing a label
The first step in tracking a virus is labeling it. To do so successfully, you will need to consider both the size of virus and what you’d like to observe. Viruses range in size from approximately 25 nm, such as picornaviruses, up to nearly a micrometer long, such as paramyxoviruses. Inserting a gene that encodes a fluorescent protein into the viral genome provides one tried-and-true way to make a virus glow. With larger viruses, such as the 400 nm vaccinia, researchers can generally insert the genes for classic fluorescent protein labels, such as green fluorescent protein (GFP) or red fluorescent protein (RFP), without interfering with the virus’s structure and function. Smaller viruses such as influenza, which averages about 100 nm in diameter, are much harder to label with expressed proteins because the size of the protein can interfere with normal viral function, Brandenburg says.
One way to tackle these smaller viruses is to use fluorescent small molecules to chemically label lipids, proteins, or carbohydrates within them. Such labeling molecules attach to particular functional groups, but they react nonspecifically and therefore don’t allow you to know exactly which molecule you are tagging. Alternatively, newer, site-specific methods allow researchers to attach a chemical label to a molecule of interest. Hidde Ploegh and his colleagues at the Whitehead Institute in Cambridge, Massachusetts, developed a site-specific labeling method that uses bacterial enzymes that recognize specific amino-acid sequences to label proteins. They are currently using the approach to examine the biogenesis of influenza virus particles. In a recent paper (PLoS Pathogens,8:e1002604, 2012), Ploegh and his colleagues describe how they engineer the five-amino-acid recognition sequence for the bacterial enzyme into two influenza envelope proteins. Using a standard peptide synthesizer, they then create a fluorescent tag appended to a short peptide sequence. Finally they treat the virus with the bacterial enzyme in the presence of the tag. The method specifically labels only the proteins with the recognition sequence, Ploegh says. It also uses an enzyme that researchers can readily produce in the lab, off-the-shelf reagents, and solid-phase peptide synthesis—tools that are available to most biochemists and cell biologists.
Another basic issue is how much fluorescent label to use. In general, Brandenburg says, it’s best to use as many copies of a label as possible without affecting the biology of the virus. Over time, light exposure photobleaches fluorescent dyes, so the more you use, the longer you can observe a virus before the dye fades. But there are lots of factors to consider. If you choose to build a fusion protein that combines your protein of interest with a fluorescent tag, Brandenburg says, make sure the labeling protein is small, and folds fast.
Different labeling techniques have distinct perks. If you’ve labeled a protein within your virus with GFP, which is expressed as the virus assembles, you could use that to follow a certain molecule of interest throughout the experiment, Ploegh says. Chemical labels, meanwhile, allow researchers to tag viruses at a particular time point, which can be useful for constructing pulse-chase experiments, where groups of molecules present on the virus at a particular point are labeled and then followed over time. Such studies allow researchers to monitor changes in those molecules during the life cycle of a virus.
It glows, but is it a virus?
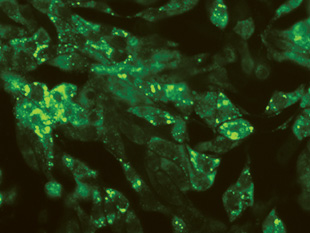
After labeling your virus of interest, you’ll need to conduct a series of preliminary experiments to make sure that the labeling worked and that you can distinguish single viral particles. Because the observable fluorescent spot is typically larger than the actual virus, you can’t immediately know whether you’re observing a single viral particle, several viruses stuck together, or a clump of fluorescently labeled aggregated protein. Simple photobleaching experiments are a first step in understanding the characteristics of your sample. If you allow a sample of your material to photobleach, you’ll observe a distribution in the rate at which the spots fade. If only labeled virus is present, those spots should bleach at a similar rate. The narrower the distribution, the more homogeneous your sample: “The majority of particles should behave similarly,” Brandenburg says.
The details of the characterization experiments may change depending on the particular virus and its biology. For example, Sanford Simon and his colleagues at Rockefeller University in New York City have used different criteria for distinguishing a fully formed virus from other fluorescent bodies in their studies of HIV than in their studies of hepatitis C. Making that call is always a challenge, as Simon’s group discovered when they first began working with HIV. “Since neither we nor anyone else had observed a single virus assemble before, it wasn’t really clear what it was that we should be looking at,” he says. “We didn’t even know whether assembly should be taking place over the course of a few seconds, or a minute or two, or tens of minutes.”
His group labeled the HIV structural protein Gag with GFP, and eventually established four criteria specific to that virus that pointed toward assembly (Nature, 454:236-40, 2008). First, they examined the distribution of photobleaching within the sample. They then looked at whether the particles they observed could restore their fluorescence: a fully formed virus would not continue to exchange with its environment and would restore its fluorescence slowly. Next, they studied the organization of proteins in the fluorescent particles. Using FRET, or Förster (fluorescence) resonance energy transfer—a technique that follows the transfer of fluorescence between closely packed molecules—they could probe whether labeled proteins were tightly organized, as they would be in a virus, or were more loosely associated. Finally, they examined the chemistry inside the viral particles. By altering pH levels in their samples and using pH-sensitive proteins to monitor changes, they could determine whether a particle had fully separated from a host cell.
You may need to consider other details specific to your virus, but Simon emphasizes that a key issue is making sure that you have clear criteria so that other researchers can replicate your experiments.
What else can you see?
Viral imaging experiments don’t just examine the structure and behavior of viruses themselves. They are also starting to offer a window onto disease processes.
For example, using labeled viruses has allowed researchers to track the movement of genes during infection. Adolfo García-Sastre and his colleagues at Mount Sinai School of Medicine in New York City used fluorescent markers to look at influenza infection patterns in mice (PNAS, 107:11531-36, 2010). Instead of labeling a structural protein, they engineered a virus that included GFP encoded in its genome so that they could track which cells were infected—a challenge, because the virus’s small size makes it difficult to label without interfering with its behavior. The modification tags a protein that affects virulence, allowing the researchers to follow its expression within the lungs of mice and identify changes in response to treatment with the antiviral drug oseltamivir.
García-Sastre sees imaging as a research tool that allows virologists to probe such questions as what types of cells become infected, what types of cells pick up antigens, where the antigens end up, and how the infection spreads.
Peeking under the surface

• MuLV (murine leukemia virus) budding from the surface of an infected mouse embryo fibroblast cell
• HIV virions budding from the surface of an infected human lymphocyte
• The 20-sided icosahedral capsid of the giant mimivirus, showing the unique structure known as a stargate, which sits atop one icosahedral vertex.
• High magnification of a facet edge of a mimivirus capsid
Fluorescence isn’t the only way to visually track a virus’s activity. If you’re interested in viral structure and in slower processes taking place in living viruses, atomic force microscopy (AFM) serves as another tool—and one that’s underused by biologists, says Alexander McPherson of the University of California, Irvine (Microbiol Mol Biol Rev, 75:268-85, 2011).AFM examines surfaces. This type of microscope consists of a nanoscale cantilever that moves along a surface—in this case, a virus. A laser shines light onto the top of the cantilever. The light bouncing off that cantilever at different angles provides an image based on the topology of the virus. To look inside the virus, you can use reagents that peel away its layers—envelope (if present), protein capsid, and nucleic acid core—one by one to acquire a series of images that reveal the virus’s structure. AFM’s resolution is approximately 10 Ångstroms, which limits its ability to examine the features of small viruses, but the approach works especially well for imaging larger ones, McPherson says, and it doesn’t require viruses to be crystallized, stained, or fixed. AFM can capture structural changes in live viruses, but only relatively slow changes. So for following processes that occur quickly, fluorescence microscopy is still the tool of choice.
A typical AFM machine costs $150,000–$200,000, and the room where it is operated needs to be quiet and free of other vibrations. The method creates images in real time as the sample is scanned. Overall, the technique is easy to learn, McPherson says, but acquiring the skills to create high-quality images takes time and experience.
Interested in reading more?


, mitochondria (red)<br >, and microfilaments (blue).")

